by Kunjumon Vadakkan
Welcome!
Objective: To understand how first-person inner sensations (in the mind) of higher brain functions (such as memory & perception) occur both independently & along with third-person observed motor activities such as speech and behavior.
Dedication: To all those who suffer from diseases of the brain (& therefore mind), especially who are abandoned by their families.

How to understand something that cannot be accessed by our sensory systems? A method used in physics
A deep principle useful for solving the nervous system - demonstrated by an example
A medication for unrelated neurological and psychiatric disorders - What does it inform us?
Insulating extracellular matrix - How thick it is? Can information get etched on it?
Has learning-mechanism got features of an evolved mechanism?
There is no need for a separate mechanism for working memory
How is learning related to LTP induction? An explanation
Extreme degeneracy of input signals in firing a neuron
Does the brain do retrograde extrapolation?
Importance of triangulation in verifying a mechanism
Testable predictions made by semblance hypothesis
Perception from a first-person frame of reference
Without sleep, there is no system! An explanation
Internal sensation - A comparison with electromagnetism
|
If you are new to this
website, then please don’t read
content in this box
first. Please read about the hypothesis and come back here.
The semblance hypothesis has put forward a model of operations of the nervous system. Here, spatial relations between specific signals arriving from input neurons are registered in “islets of inter-LINKed spine heads (IILSHs)”, whose spines belong to different output neurons. In response to a prompt (cue stimulus), operation of IILSHs generates both first-person inner sensations and motor outputs such as speech and behavior. The field of artificial intelligence uses artificial neural network where neurons are connected with each other in different configurations. The configuration in large language models (LLMs) has succeeded in showing features of good generalization - ability of a trained system (nervous or artificial) to perform well on new input data unseen during training. This prompted to examine the basic concept of operation in the LLMs. This shows that "attention heads" within the hidden layer located in between input and output neuronal layers of Transformer in LLMs is equivalent to a linear algebraic treatment of one segment of operation of "islets of inter-LINKed spines" that generates motor outputs. Article
If a primary school child ask me, I will tell this work as a STORY |
|
This is a video uploaded on 8th July, 2023. If you are new to this website, then please read this front (home) page and FAQ section of this site before watching. An explanation for associative learning in terms of semblance hypothesis
Video (Note: Qualia is a virtual first-person property. Qualia of inner sensation can be estimated by a retrograde extrapolation from postsynaptic terminal D towards the sensory receptors. Even though it is shown as part of the video for demonstration purpose, there is no back propagation of potentials in that direction. Generation of inner sensation is a system property of systems that has oscillating extracellular potentials whose vector components are contributed by synaptic transmission across synapse A-B & propagation of potentials along the inter-postsynaptic functional LINK B-D in a perpendicular direction to the synaptic transmission). This video provides an explanation for changes that occur when sound & foot shock are associatively learned. It then explains how sound alone (after learning) triggers inner sensation of memory of foot shock & cause foot withdrawal. Neutral sound is the conditioned stimulus (CS). Aversive foot shock is the unconditioned stimulus (US). Both reaches neurons of Lateral Amygdala (LA). Certain learning-change is expected to occur at locations of their convergence in LA. After associative learning between sound & foot shock, sound alone causes an inner sensation of foot shock & triggers foot withdrawal. A testable explanation for a link between pathways of sound & foot shock where they converge in LA is expected. At the location of convergence, sound should generate inner sensation of foot shock (in the absence of foot shock). This is like a sound-induced hallucination of foot shock. Marvin Minsky gives us some guidance - onlinelibrary.wiley.com/doi/abs/10.1207/s15516709cog0402_1
Questions are a) "Is there a suitable location where
hallucinations can be triggered by sound during memory
retrieval?" b) “What are the pre-conditions that set a
stage to trigger hallucination of foot shock when sound
arrives?”
This can be explained using two examples. a) First is a
method by which a
veterinarian gives an intravenous injection to a horse.
Initial
patting on the side of neck (over the visible internal
jugular vein) followed by tapping with increasing intensity
enables the veterinarian to rapidly
place a needle in between the fingers of the tapping
hand to get inserted to that vein without notice. b)
Second is about pickpocketing. It
is easy to get pickpocketed while climbing stairs
(e.g., in a railway station) since the
gluteal region is frequently moving while taking steps. From both, it is
possible to insert/remove something to/from a system
that is undergoing a continuous sequence of events without
the system’s notice.
In both cases, newly inserted stimulus is perceived as
one of the frequently occurring events. i. e., the newly
inserted stimulus leads to a hallucination (at the
location of insertion) that it is receiving a usual
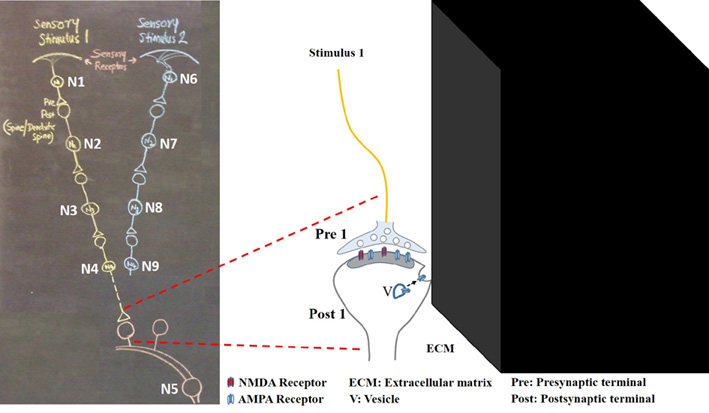
stimulus that has been occurring prior to it. A postsynaptic terminal (spine) is continuously being depolarized by quantally released neurotransmitter molecules (NTMs) from its presynaptic terminal. When an action potential arrives, the latter releases a volley of NTMs to trigger a large postsynaptic potential. For a postsynaptic terminal (spine), it receives inputs from its presynaptic terminal all the time & sometimes from the environment via its presynaptic terminal (until learning occurs). This is the dominant state of the system. In the above contexts, if learning cause formation of an electrical link between spines (that belong to different neurons, since spines on a dendrite are separated (mean value of inter-spine distance) from one another by a distance more than the mean spine diameter) to which associating stimuli arrive, then we can examine whether it will facilitate a mechanism for memory retrieval. Later, arrival of one of the associatively learned stimuli through the link that reaches the lateral aspect of the spine (and depolarizes the entire spine) to which the second stimuli had arrived during learning will be perceived as if the second stimulus has arrived from its presynaptic terminal. This forms the basis for a hallucination (as Marvin Minsky envisioned). In other words, depolarization of a spine from its lateral aspect tricks the latter to hallucinate that it receives an input from its presynaptic terminal (from the environment).
Since mean inter-spine distance is more than mean spine diameter https://pubmed.ncbi.nlm.nih.gov/12838576/, the above link is expected to occur between spines that belong to different LA neurons. The potentials elicited by sound propagates through the link to reach the postsynaptic terminal through which shock arrived in the past to trigger hallucination (inner sensation) of shock. Potentials then propagate further to trigger foot withdrawal. There should be a mechanism to integrate units of inner sensations originating at many nodes of convergence to cause inner sensation of foot shock. The brain functions only in a narrow range of oscillating extracellular potentials. A cue stimulus reaching the link leads to propagation of depolarization in perpendicular directions - through synapse A-B (see video) and link B-D. These can provide vector components to oscillating extracellular potentials. Thus, we have a mechanistic explanation for a testable change that occurs during associative learning & a mechanism for cue-induced cue-specific hallucination that generates inner sensation of foot shock & foot withdrawal.
|
|
If you are new to this website, then please don’t read
content in this box
first. Please read about the hypothesis and come back here.
A new explanation for
flash-lag effect using a testable mechanism for
first-person inner sensation of perception When a flash is briefly presented in a specific location adjacent to the path of a uniformly moving object, the former is perceived to lag the latter. Different experimental conditions carried out to qualify this flash-lag effect (FLE) have led to several seemingly unrelated first-person reports from which several constraints are available to synthesize a testable mechanism for FLE. Using a derived mechanism for the generation of first-person internal sensation of perception, present work provides new interconnected explanations for disparate findings associated with FLE. By verifying testable predictions arising from the hypothesized mechanism of first-person inner sensations, it may become possible to understand timescale-matched neurobiological changes responsible for perception. Article |
|
If you
are new to this website, please don’t read
content in this box
first. Please read about the hypothesis and come back here. Golgi staining of neurons: Oxidation state-dependent spread of chemical reaction identifies a testable property of the connectome Abstract: Camillo Golgi observed the reticular nature of the nervous system by his staining method. Ramón y Cajal modified this protocol and obtained staining restricted to individual neurons, which was in agreement with the cell theory. Close examination shows that Golgi used an oxidizing agent to pre-treat the brain tissue before the staining reaction and Cajal used an additional oxidizing agent for the same step. It shows that oxidation state of the tissue has a crucial role in determining the spread of Golgi chemical reaction between neurons. Since a) dye injected into a neuron spread only within that neuron's cytoplasm, and b) it is possible to grow individual neurons in primary culture, oxidation state-dependent spread of chemical reaction should take place through a gate while keeping the cell membrane intact. These constraining features guide towards the solution where formation of such a gate between spines of different neurons is a suitable mechanism that can take place during learning. As long as this gate exists, it should allow propagation of depolarization from a cue stimulus across the gate and provides options to a) generate internal sensation of associatively-learned item, and/or b) provide potentials to cross the threshold to fire neurons to generate motor action reminiscent of the arrival of associatively-learned second item. These constraints lead to the derivation of a crucial connection that can define the connectome. This oxidation state-dependent reversible connection of the connectome matches with the operational functions of inter-neuronal inter-spine functional LINKs proposed by semblance hypothesis. All the above findings are testable. Article |
A second view of the problem can be described as follows. It begins by examining the following situation: The heart pumps the blood, and the kidneys filter the waste materials from the blood; these functions are observed by us from a third-person perspective. We understood their functions quite well, evidenced by our ability to develop their replacements, such as artificial heart and dialysis. What functions does the nervous system carry out? What operational mechanism do we need to understand from brain to replicate/replace it? Brain generates an inner sense of the external world during perception, stores sensory information during associative learning and later produces the internal sensation of retrieved memories of the learned item when the associatively learned cue stimulus arrives, induces thought process to connect different items from different sets of learning events – all of which are first-person properties that cannot be accessed by third-person observers. The only information that are available from the owner of the nervous system to a third-person are from the surrogate markers consisting of motor activity - specifically, speech and behavior. Therefore, the pieces of the puzzle mentioned in the above paragraph should be capable of explaining both first- and third-person accessible functions.
A third
view of
the problem
becomes evident when examined from the
viewpoint of a builder. Here, the job is to replicate the nervous
system in an engineered system.
A fourth view becomes possible by observing the “loss of function” states of the system occurring at various levels. This can help understand the nature of the pieces of the puzzle. Early genetics research has gained valuable information from "inborn errors" of metabolism that provided guidance to understand the allelic organization of genes. In this context, examining neurological and psychiatric disorders can help to understand the nature of the operating mechanism. Since the exact pathological features of many of these diseases are not yet known, it is expected that the loss of function of the operational units that induce both the first-person and third-person features is expected to provide information about the pathologies from which the function can be verified.
A large number of features observed by different subfields of neuroscience and psychology are required to be explained by a solution for the system. Since these features are very diverse, only a unique cellular mechanism will be able to explain all of them. This is expected to be a unique structure-function mechanism occurring at the intersection between the third person observed features and first-person properties. In other words, it is a dynamic, but stabilizable structural feature that can provide basic units of first-person internal sensations of different higher brain functions. It is necessary to verify whether the derived solution can explain findings from different specialized faculties within the large fields of neuroscience and psychology and test whether the explanations are inter-connectable. Since our current research efforts in each field are moving towards more specialized and super-specialized areas, finding and verifying the unique solution (to put the pieces of the puzzle together) requires an effort in the opposite direction. Anticipating this is the most important requirement for solving the system. It is necessary to explain how the nervous system functions occurring at different levels, such as - a mechanism that directs potentials to induce the internal sensation concurrent with the activation of motor neurons at physiological timescales (interconnecting central mechanism), dendritic spine changes, long-term potentiation (LTP), place cell firing, consolidation of memory, and association of memory with a feasible framework for consciousness - are interconnected.
For arriving at an operational mechanism that can explain both third person and first-person properties, a theoretical approach is the most efficient method. Among different brain functions, memory has the advantage that experiments can be carried out both to associatively teach the system and examine how these learning-induced changes are used during memory retrieval. A very large amount of experimental data is available in the field of memory research. Since no cellular changes are observed during memory retrieval, the memory retrieval is likely taking place by a passive reactivation of a learning-induced change. Memories were classified into working, short-term and long-term based on the differences in the period of time, following learning, during which they can be retrieved. Studies have been carried out with the assumption that the cellular mechanisms during learning that leads to memories classified in this manner are different. Since qualia (virtual first-person internal sensations) of these retrieved memories are almost same, it prompts one to ask, "What if a) a common cellular mechanism is taking place during learning, and b) the retrieval of different types of memories can be explained by reactivation of learning-induced changes that are retained for different durations?" To undertake such an experimental approach, one may ask, "Can we directly examine the memories themselves instead of examining the motor activity such as behavior and speech at the time of memory retrieval?" This will also eliminate examination of slow molecular changes following learning. In this context, it is necessary to re-define the question: "What are memories?" Memories are first-person virtual internal sensations of an item (in the absence of that item) induced within the nervous system (in response to a cue stimulus or occurring spontaneously). The sensation of a stimulus in its absence is hallucination. Therefore, memories can be viewed as cue-induced cue-specific hallucinations. Can we search for a learning-mechanism that can allow induction of virtual first-person internal sensations of memory as a cue-induced hallucination? This is the basis of developing semblance hypothesis which was published first as a book in 2007 (a copy is uploaded in Publications section). Revised editions were published in 2008 and 2010.
There are three main reasons why we had difficulties in solving the nervous system.
1) Frame of reference problem: Every time we had a frame of reference problem in the past, we needed to pause for some time to make further progress. A typical example is the difficulties in sensing the rotation of Earth (note that the speed of rotation of Earth on its axis is 1670 km per hour). Our sensory systems cannot sense rotation of Earth towards the East since we are in the same frame of reference as that of the Earth. The fact that third-person observers cannot sense first-person inner sensations in a subject can be viewed as a frame of reference problems. Physics has developed methods to solve frame of reference problem. In the case of the nervous system, we need to use the principles of methods used in physics to overcome this hurdle to successfully solve it. Since this is new for neuroscience, there will be some difficulties in thinking about it in the beginning. But hopefully we will soon understand the problem and accept methods to overcome it.
2) Difficulty to study the “virtual”: Another difficulty is the virtual nature of first-person inner sensations. However, we have had the experience of dealing with virtual items in the past. For example, numbers do not exist. We made them. In fact, they are virtual in nature. We can say that they represent real counts of items. What about negative numbers (integers?). They can exist only in our imagination. Yet, we use them routinely in mathematics. On a graph, we don’t feel their virtual nature at all. Going one step further, we have invented complex number (imaginary number). This solved our difficulties in finding square roots of negative integers, which helped to further advance mathematics. In a similar manner, once we understand where and how units of inner sensations are sparked within the system, we will be able to perceive a virtual space where we can position them and navigate that space to understand their different conformations.
3) Access problem: How to understand something that cannot be accessed by our sensory systems? There are several examples where we succeeded in overcoming such problems. For example, we cannot see DNA inside the cells or in a gel. But we stain it with ethidium bromide that will allow us to see the stain through our eyes. This means that our access problem can be overcome by adding one step that allows us to use our sensory system (here, our eyes). We sometimes go two steps. For example, we use a primary antibody to an antigen, followed by a secondary antibody with florescent property that can then be visualized through our eyes. Even though our sensory systems do not have direct access, we believe that an antigen is present based on logic that we use in our staining protocol. Understanding inner sensations will need another such indirect method that we will eventually become familiar with .
How did Galileo show
us to put together observations to make an inference even when our
sensory systems cannot agree with such an inference?
When are many observables are present in a
system, putting them together can provide a totally new inference
that our sensory systems may not be able to directly sense. So,
naturally such type of an inference often will not be accepted by
our community quickly. But every scientist has learned how to uphold
the importance of this scientific method. An often-cited example is
that of the inference made by Galileo Galilei using different observations
that he made using his new telescope. Galileo published his findings
in his book “The starry messenger.” While observing Jupiter on 7th
January 1610, Galileo found four moons that are orbiting Jupiter.
He immediately made the conclusion that if these moons are revolving
around Jupiter, then it is unlikely for the Earth to be at the
center of the Universe. A video explains this.
www.youtube.com/watch?v=NXOYqTUpkaM
Galileo then turned
his telescope towards Venus. It showed phases similar to that of
Moon - New Moon and Full Moon. Galileo observed both the New and
Full faces of Jupiter. When it is New, it is very big. When it is
Full, it is very small. Galileo concluded that this could happen only if Jupiter
revolves around the Sun and therefore, he made the inference that
Sun must be at the center of the system. A suitable fit with this
observation is that Earth and other planets are also revolving
around the Sun. What we now know is that Jupiter is New when it
comes close to the Earth, blocking the light from the Sun.
When Jupiter is on the other side of the Sun during its
revolution around the Sun, it is farthest from the Earth. Here,
Jupiter is seen as small, and its face is Full. A nice video explains
it is here.
www.youtube.com/watch?v=W-6x3XRuWVg
The above two observations made Galileo conclude that Earth is not at the center of the solar system. Galileo saw the simplicity if all planets revolved around the Sun. Galileo was making logical arguments that allowed him to fit all the findings together. We have such a rich tradition in science to gather information from different observations and then put them together to make an inference even though we cannot appreciate it directly with our sensory systems. Another limitation of our sensory systems is their inability to sense the speed of rotation of the Earth on its axis. Since the circumference of the Earth at the equation is nearly 40,000 kilometers and one day has 24 hours, we can make the inference that the speed of rotation of the Earth is nearly 1650 kilometers per hour. Even though we cannot sense this speed using our sensory systems while on Earth, our inference about the speed of rotation of the Earth (and ours) must be true. When we observe the Earth from space, we see rotation of the Earth. See NASA’s video from the international space station (ISS) located 408 kilometers from Earth. https://www.youtube.com/watch?v=86YLFOog4GM (This time-lapse video from ISS needs to be converted to real-time video).
In short, we must find ways to overcome the limitations of our sensory systems! So, the question is, "How can we understand the operation of the nervous system even without replicating the mechanism in engineered systems?" In the case of the nervous system, our job is to put all the observations together to make an inference that will interconnect all those observations. It is obvious that the main function of the nervous system is generation of inner sensations as a first-person property, which a third person observer cannot sense directly. We have been thinking that it is the most difficult function to understand. However, we must obtain motivation from the above examples of finding solutions for non-sensible events. We must be willing to derive a testable solution using observations from multiple levels of the system and put forward testable predictions that we can verify.
We need to bring all
the observations of the nervous system together and make an
inference (solution). In this exercise, we cannot afford to leave
out even a single observation since we must make sure that we
reach a solution that can interconnect all the features of the
system. When we become able to derive a
solution that can provide explanations for all the functions, then
we can make a reasonable assumption that the derived solution is correct. We must use this solution to make
testable predictions that we can go and verify.
Footnotes:
It
was Copernicus, who based on observations, first proposed that the
Sun is stationary and that the Earth and other planets revolve
around it. So, the credit must go to him as the first person who
reported that Earth is revolving around the Sun. Even
though Gallio thought that planets are moving in circles, Kepler
found about the elliptic path of motion of planets.
How can we solve the nervous system?
As we face situations that have more steps away from reality, we have to rely on our logical reasoning capabilities (see an excerpt from Krakauer et al., 2017 that appeared in the journal Neuron). We have a very large number of findings from different levels, and we have to discover the solution that can interconnect all of them. We need to overcome a) the frame of reference problem, b) the access problem, and c) to deal with the virtual nature of internal sensations. But what if we cannot sense one part of the solution directly by our sensory systems while trying to find the solution? In this case, it is possible to seek examples of approaches that are used by other fields of sciences. For example, physics study particles and fields that are not accessible to our sensory systems. What is the deep underlying principle behind their success? A summary is given in Table 1 below.
|
Physics |
|
| 1) First, a large number of observations are made that appear to be disparate in nature. This means that these findings cannot be explained in terms of each other. | 1) There are a large number of disparate findings in neuroscience (see Table 2) that need inter-connectable explanations. Example: How does the operation of the system relate to sleep and also to the electrophysiological finding of LTP? |
| 2) The above indicates the presence of a deep underlying principle that should interconnect these disparate observations. | 2) There should be a deep underlying principle behind all those observations in Table 2. |
| 3) The effects of the above principle are the ones (e.g. particles and fields) that cannot be directly sensed by our sensory systems. | 3) There is a principle, the products of which (inner sensations) cannot be sensed by our (third-persons') sensory systems. Yet, the principle of the mechanism should be able to explain and interconnect all the observed findings. |
| 4) The next step is to search for any possible solution that can interconnect all the findings. Constraints provided by disparate observations should be able to guide us towards the solution. This is done either by initial deduction followed by mathematical approximations (Special and General Relativity) or by pure mathematical derivation (Higgs Bosons). | 4) A structure-function mechanism has to be sought by logical deduction & trial and error methods. All the constraints offered by a large number of findings can be used to derive the solution. Success depends on moving through a path as guided by all the constraints. Only when we reach the correct solution, we will be able to explain all the findings in an interconnectable manner. |
| 5) The solution is then confirmed by verifying the testable predictions. | 5) Testable predictions made by the derived mechanism can be verified. |
Table 1. Comparison
between the steps
taken by physics and neuroscience when it becomes necessary to unify
disparate findings. This is important especially when dealing with
properties that are
non-accessible to our sensory systems such as particles, fields, and
first-person internal sensations etc.
The deep underlying principle of many studies in physics has similarities to the method used in linear algebra for solving a system of large set of linear equations that has a unique solution. If one tries to solve such a system, one can find that the relationships between the variables in each equation provide hints that can guide us towards the solution. If there are a large number of variables, there should be at least an equal number of equations to find the unique solution. Since there is a large number of findings at different levels of the nervous system, we can (and we must) use even minor interconnected features between them to find the solution. It is a gigantic exercise since there are no easy methods in biology like that are used in linear algebra. But understanding linear algebraic methods will definitely help in solving the system.
In linear algebra, Gauss-Jordan elimination method is used to find the solution for a system of linear equations. If we look carefully, we can see that easy methods in linear algebra were designed by someone who understood the deep underlying principle and worked on to make it simple for others by developing easy methods. We can examine how the relationships between variables in each equation define the unique solution for a system of linear equations and how Gauss-Jordan elimination method was invented. It is to be noted that we can also solve linear algebra problems using trial and error methods. But it will take some extra time. In other words, in mathematics easy methods are developed for convenience. Whichever method is used, the deep underlying principle is the same - A system exhibiting a large number of disparate findings (equations) most likely has a unique solution that binds (interconnects) all the findings (equations) within that system. By finding a solution that can interconnect a subset of findings and by repeating this approach using different subsets of findings, one can hope to reach a common underlying solution, which is the correct solution. One can start attempting to solve the (nervous) system by using subsets of disparate findings. The optimism with this approach is that there is only one unique solution for the nervous system, and it is easy to rule out many wrong solutions quickly. Using the above principle, subsets of constraints provided by findings from various levels (Table 2) were used to derive a mechanism that can explain, and interconnect findings made by different faculties of brain research. This approach is expected to lead to the derivation of an operational mechanism for the generation of internal sensations, of which the non-sensible component will continue to remain non-sensible to our sensory systems even after its discovery (Figure 2). However, it is expected to show testable learning-generated change that gets reactivated at the time of memory retrieval to induce basic units of internal sensation whose integrational product can provide sensory qualia of the retrieved memory. Structural and electrophysiological changes that are expected to occur from these changes are expected to get explained using experimental results from different laboratories.
There are very large number of observations from different levels of the nervous system functions (Table 2). An example of store receipts (this is also the deep principle behind some of the approaches carried out in physical sciences to understand Nature) provides us confirmatory evidence that we can reach a final correct solution, if it becomes possible to obtain large number of findings that contain all the variables of the system. From a philosophical standpoint (Excerpts from an article here) also, we need to undertake a similar approach by examining findings from all levels together. This approach is expected to provide information about a testable mechanism that generates first-person inner sensations.
|
Nonsensible (not
directly with our senses) features of a
system can be inferred from its sensible properties. Since
we have a large number of constraints from findings in multiple levels of the nervous system, a
derived solution that can explain all its features
at those levels is
likely correct even if we are unable to directly sense the
formation of first-person inner sensation. Further, we can
a) do retrodictive
examinations, & b) make testable predictions
for verification. |
|||
|
|
The findings |
Constraints offered by the findings (on the left side) that
direct the enquiry towards a correct solution.
|
Interconnected explanations by the semblance hypothesis
(Please read this row after reading the hypothesis) |
|
1 |
Both associatively learned stimuli &
prompt (cue) stimuli propagate through synaptically-connected neuronal
circuit. |
Mechanism should operate synchronous with the
synaptically-connected circuitry |
Inter-neuronal inter-postsynaptic functional LINKs (IPLs)
form and
operate only when synaptic transmission takes place (Vadakkan,
2007;
2013) |
|
2 |
Learning-induced changes occur in
physiological timescales (in milliseconds)Foot note1 |
A learning-inducible change that
occurs (& completed) in physiological timescales (to explain
the ability to retrieve memory instantly following learning) |
Propagation of potentials along the IPLs to the inter-LINKed
spines takes place in physiological timescales of
milliseconds (Vadakkan, 2007;
2013) |
|
3 |
Memories that can be retrieved after
long period after learning are also capable of getting
retrieved immediately following learning (working memory) |
Learning
should generate retrieval-efficient changes within
milliseconds. They can be used for memory retrieval
immediately (working memory). These changes have a provision
for remaining in a stable form for long period,
responsible for long-term memory |
IPL
formation takes place at the time of learning. When IPL
reverses back, then its short duration can only generate
working memory. The life span of IPL decides the duration of storage
of memory. Long-term stabilization of IPLs lead to long-term
memory
(Vadakkan, 2010a;
2013) |
|
4 |
When exposed to a cue stimulus, inner sensation of memory occurs
in physiological timescales (in milliseconds) |
A
learning-induced change should be capable of inducing inner
sensation of memory
in physiological timescales (completed
within this time). |
Propagation of potentials along the IPL generates semblance
over the inter-LINKed spine instantly as a system property
(Vadakkan, 2013) |
|
5 |
Memory is an internal sensation of an
item/event having certain specific sensory features (qualia) |
Mechanism
is expected to have elements that
can provide sensory features to the retrieved memory.
|
Integration of all the units of inner sensation generated on large number of inter-LINKed spines by a cue stimulus
provide qualia (Vadakkan, 2010a) |
|
6 |
Ability to store large set of
learning-induced changes responsible for retrieving large number
of memories |
Neurons and
their processes are finite in number. Therefore, an
efficient operation for storing large numbers of
learning-induced changes becomes possible if common elements
in each learning mechanism can be shared. Hence, each memory
is expected to get induced from a combination of unitary
mechanisms.
|
Inter-LINKed spines within the islets of inter-LINKed spines
can be depolarized by any specific stimulus reaching it.
Since items/events consist of combinations of sensory
stimuli, different combinations of inter-LINKed spines can
be used by different cue stimuli to generate corresponding
memories (Vadakkan, 2010a) |
|
7 |
Instant access to very large memory stores. |
A specific cue stimulus should be able
to induce a specific memory by combinatorial reactivation of
a specific set of learning-induced unitary changes |
Inter-LINKed spines can be accessed by stimuli
from any cue stimulus (Vadakkan, 2010a) |
|
|
Absence of cellular changes during memory
retrieval.
|
A passive reactivation of the changes that occur during learning should be getting used at the time
of memory retrieval to induce units of internal
sensations. This should take place at physiological
timescales of milliseconds.
|
Since inter-LINKed spines
persist from the time of learning,
propagatoin
of depolarization
along the IPL does not require any new cellular changes (Vadakkan, 2010a) |
|
9 |
During memory retrieval, firing of a subset of neurons that were not firing before learning in
response to the same cue stimulus |
Learning has opened certain new
channels & cue stimulus leads to propagation of
depolarization through these channels to provide additional
potentials to a subset of neurons that are otherwise being
held at subthreshold activation state (without firing). This
will lead to firing of these set of neurons. |
Formation
of IPLs during learning will lead to propagation of
depolarization across them to the inter-LINKed spines. This
will provide additional potentials to the inter-LINKed
spine’s neurons and may fire those neurons (Vadakkan, 2013) |
|
|
Brain operates
in a
narrow range of frequency of extracellularly recorded oscillating potentials. |
Expected mechanism
provides vector
components
of the
oscillating potentials.
|
Synaptic transmission & near perpendicular propagation of depolarization
across the IPL provides vector components. Reactivation of a
set of IPLs at rest generates a background semblance, which
is
expected to generate inner sensation of a conscious state (Vadakkan, 2010b) |
|
|
Motivation promotes learning. Motivation is associated with
release of dopamine at different locations of the brain |
Motivation is associated with specific factors
and their specific actions are expected to promote the
learning-induced change and possibly to retain this change
for longer period than that occur in its absence |
Dopamine is known to cause
spine expansion (Yagishita et al., 2014). Expanding spines can augment IPL formation and retain the formed
IPLs for long time that may trigger some stabilization steps |
|
|
Internal sensations of working, short,
and long-term memories have similar
qualia |
The same
learning-induced change is retained for different durations.
Long-term memory loses its clarity both due to loss of some
unitary mechanisms & dilution of specificity by combining
with newly formed units of inner sensations.
|
Memories of all durations take
place by reactivation of inter-LINKed spines by
depolarization propagating along the IPLs and generating
units of inner sensations (Vadakkan, 2010a,
2013) |
|
13 |
Working memory lasts only for a very
short period of time |
Learning-induced change must have a
quickly reversible mechanism. |
IPL
formation is a high energy requiring process as can be
inferred from experiments using artificial membranes (Rand and Parsegian, 1984;
Martens and McMahon, 2008;
Harrison, 2015). Hence majority of IPLs are
expected to reverse quickly |
|
14 |
Some of the same memories that are
retrieved as working memories can be retrieved after a long
period of time
after the learning. |
Learning-induced change must be able to undergo certain changes that will
enable it to get maintained for a long period |
Stabilization of IPLs for long period can induce the same
units of inner sensation. If the number of IPLs that can be
reactivated by a cue stimulus after a long period decreases,
then qualia of memory will reduce (Vadakkan, 2010a;
2013) |
|
15 |
Simultaneous existence of previous
two conditions (above two rows) within the system |
Learning-induced mechanism should have
an initial quickly reversible change that under certain
circumstances can progress towards a stage where it can get
stabilized for long period of time |
When
memory of a beneficial or deleterious item or event becomes
advantageous for survival, then IPLs necessary for those
memories are stabilized for long period (Vadakkan, 2010a;
2013) |
|
16 |
Internal sensation of memory
in response to a
cue stimulus varies with the nature of
the cue stimulus |
Specific sensory features from the cue stimulus induce a
specific combination of internal sensory units to generate
specific features of memory being retrieved.
|
As the cue stimulus propagates through its path, depolarization
propagates through a specific set of IPLs that induce a
specific set of units of inner sensations for a specific
memory (Vadakkan, 2010a,
2013) |
|
17 |
After associative learning between
two items, arrival of one of the items generates memory of
the other item |
The learning mechanism should have
features to explain how either one of the associatively
learned items can act as a cue stimulus to generate memory
of the other item. Hence, the mechanism should have the
ability to show bidirectionality in it |
Semblance
can be generated from either one of the inter-LINKed spines
on the two sides of an IPL (Vadakkan, 2010a;
2013). |
|
18 |
Even partial features of one of the
associatively learned item is capable of retrieving memory
of the second item |
The mechanism should have features to
explain how stimuli from partial features of one stimulus
can retrieve memory of the second item. |
Partial
stimuli propagate to generate inner sensation of a framework
of an item or event. Due to the property of generalization
(by virtue of spread of depolarization across the entire
islet of inter-LINKed spines), it is expected to provide
more features to the memory (Vadakkan, 2010a;
2013;
2019a) |
|
19 |
Ability to store new memories without needing to overwrite the old ones |
Sharing of unitary mechanism for
common features, and provision for formation of new units
with new associations are expected to be present in the
system |
Inter-LINKed spines within islets of
inter-LINKed spines can be shared by any stimuli reaching
them. Hence there is no need for overwriting old memories.
Any few associations can get inter-LINKed to the existing
islets of inter-LINKed spines
(Vadakkan, 2010a;
2013) |
|
|
Consolidation of memory (Transfer of storage locations of
memory from the hippocampus to the cortex) over a span of 5
to 8 years |
Addition of specific
learning-induced changes in the cortex over time using
similar unitary sensory associations, and ability to
generate memories by a global integrating mechanism. Must go
through a stage of surplus unitary mechanisms |
Convergence of all sensory
stimuli in the hippocampus leads to dense islets where
inter-LINKed spines can be formed. Sparser IPLs are expected
to be formed in the cortex. Neurogenesis and repetition of
associative elements within each learning will lead to
formation of surplus IPLs in the cortex over time
(Vadakkan, 2011a). |
|
21 |
Mechanism uses pre-existing schemas (Tse et al., 2007). Schemas
are expected to get used inter-changeably. |
Changes
induced by one learning are shared by another learning
event. For this to occur, there must be shared unitary
mechanisms in each learning event and presumably also in the
memory retrieval mechanism.
|
Inter-LINKed spines can be used by any cue
stimuli, allowing the unitary structural operations to get
shared. It is also reasonable to assume that units of inner
sensations induced from these inter-LINKed spines also get
shared by similar elements in stimuli
(Vadakkan, 2010a;
2013) |
|
22 |
A constantly adapting dynamic circuit
mechanism is expected |
Provisions should be present to
accommodate large number of new learning events. |
Since
extracellular matrix space is minimal in the cortex, large
number of spines from different neurons are expected to
remain abutted to each other. They can be readily
inter-LINKed depending on the nature of cue stimuli (Vadakkan, 2010a;
2013) |
|
23 |
Framework of a mechanism that can generate hypothesis by the system |
When one of the elementary mechanisms of one
associative learning event undergo association with a third
elementary mechanism during a third associative learning, it
will lead to an interconnected chain of associations (ICAs).
When there are common elements in different ICAs, then the
system will be able to generate a hypothesis of
relationships between events/items.
|
When one spine each from two islets of
inter-LINKed spines are inter-LINKed, any one spine from one
islet establishes a relationship with any one spine in the
second islet of inter-LINKed spines. When more than one
islet of inter-LINKed spines is inter-LINKed in this manner,
it leads to generation of hypothesis about something that
will not occur ordinarily
(Vadakkan, 2010a;
2013) |
|
|
System needs
a
state of sleep for nearly one
third of its operational
time |
It is necessary to explain why the system won't
be able to exist without sleep. i.e. Explain substantive
nature of sleep in the operation of the system. |
State of sleep is needed to
keep postsynaptic depolarization by presynaptic terminal as
a dominant state of the system. Only when this dominant
state is maintained, then only a lateral activation of the
postsynaptic terminal (spine) will induce units of inner
sensation of a stimulus arriving from the environment
through the presynaptic terminal (Vadakkan, 2016b). Since nothing comes from the environment, it is a hallucination (Minsky, 1980) |
|
25 |
While living in space, requirement of
sleep reduces by more than one hour |
Provide a mechanistic explanation why
reduced sensory stimuli in space reduces the need for sleep.
|
Since
sensory stimuli is less in space, number of reactivations of
inter-LINKed spines is reduced. This reduces the time to set
the system to its baseline dominant state of postsynaptic
depolarization by its presynaptic terminal
(Vadakkan,
2016b) |
|
26 |
During memory retrieval, inner
sensation of memory can occur with or without motor actions
such as speech or behavioral motor actions |
The mechanism that generates inner
sensation of memory should have a connection with the
mechanism that generates motor action. There should be a
provision for disabling this connection at will |
IPL
mechanism can generate both units of first-person inner
sensations and motor action reminiscent of arrival of the
item whose memory is retrieved. The motor outputs can be
inhibited while inner sensation is being generated
(Vadakkan, 2010a;
2013). |
|
27 |
It is difficult to inhibit a memory
which is being retrieved |
A structural mechanistic explanation
is needed |
IPL is an
inter-membrane connection. Once IPL is present and
functions, it is not possible to inhibit its function
voluntarily. Additional inter-spine LINK with an inhibitory
spine may become possible through future associative
learning events (Vadakkan, 2007;
2010a) |
|
28 |
Mean inter-spine distance on the
dendrite of a pyramidal neuron is more than the mean spine
diameter
(Konur
et al., 2003) |
This opens the possibility for
neuronal processes that belong to other neurons to occupy
the inter-spine space. It is reasonable to expect some
functional importance for such a scheme of inter-spine
spacing. Since spines of different neurons occupy this space
and ECM is often negligible (see Fig.13 in FAQ), some of the
spines that belong to different neurons can remain abutted
to each other. |
Abutted
spines that belong to different neurons increase the
probability for inter-neuronal inter-spine interactions.
These interactions are the basis of IPL proposed by the
semblance hypothesis
(Vadakkan, 2010a;
2013) |
|
29 |
Both learning and retrieval of memory
are associated with firing of a set of neurons. |
Both learning and memory retrieval allow potentials
to propagate through certain gateways that allow certain
neurons that are being held at sub-threshold activation
states to fire action potentials. This gateway may be formed
at the time of learning and are likely associated with
generation of units of inner sensation of memory.
|
Both
learning and memory retrieval allow propagation of
potentials along the IPL. This leads to the arrival of more
potentials to the axon hillock of the postsynaptic neuron.
If arrival of these additional potentials can allow the
subthreshold activation state of the neuron to cross the
threshold for firing, it fires an action potential
(Vadakkan, 2010a;
2013) |
|
30 |
Place cells fire in response to specific spatial stimulus. |
Mechanism that generates inner
sensation of
memory for a location is expected to have a mechanistic
connection with firing of a
set of
CA1 neurons. |
Place cells are CA1 pyramidal cells. When islets of inter-LINKed spines
of overlapping dendrites of CA1 neurons receive spatial
inputs, they provide potentials to their postsynaptic CA1
neurons. If these CA1 neurons are being held at subthreshold
activation states, then they fire. This explains place cell
firing
(Vadakkan,
2013;
2016a) |
|
31 |
Firing of an ensemble of neurons during a higher
brain function. |
Inner sensation
generated during a higher brain function is related with
firing of an ensemble of neurons. |
Reactivation of IPLs during a higher brain
function will add potentials from the inter-LINKed spines to
their postsynaptic neurons. If these potentials add up to
allow these neurons to cross the threshold, they will fire
(Vadakkan, 2010a;
2016a) |
|
32 |
Firing of
separate sets of
neurons during learning and memory retrieval |
Learning and generation of
inner
sensation of memory are associated with firing of
separate
sets of neurons. |
When memory retrieval is immediately following learning, then lack of
stimuli from the item whose memory is being retrieved is
responsible for the difference. When memory is retrieved
late, other associative learning events in between the
learning and memory retrieval under examination will
generate additional IPLs in the circuitry & will be
responsible for the difference
(Vadakkan, 2010a;
2016a) |
|
33 |
Fast changes in both the magnitude and correlational structure of cortical network activity
(Benisty et
al., 2024) |
Rapidly time-varying functional connectivity is
responsible for such changes. |
Changes in environmental stimuli, self-triggered thought processes,
inner sensations of fear, anticipation, hunger, and comfort
levels fluctuate moment to moment indicating reactivation of
a new set of IPLs. This will change network activity (Vadakkan,
2019a) |
|
34 |
Firing of a cortical neuron (axonal spike) is possible by summation of
nearly 140 postsynaptic potentials (input signals) arriving
from random locations. Each of these cortical neurons have
tens of thousands of dendritic spines where postsynaptic
potentials get generated |
These neurons must be maintained at a
sub-threshold state in the background state and the
mechanism of induction of internal sensation must be
associated with providing additional postsynaptic potentials
for crossing the threshold for firing of these neurons. |
Recent modelling studies have shown that a pyramidal neuron that has
tens of thousands of input connections can fire an action
potential by spatial summation (summation at the same time)
of nearly 140 EPSPs at the axonal hillock that arrives from
randomly located dendritic spines (Palmer et
al. 2014;
Eyal et al.,
2018). Based on calculations of energy per bit of
information 2000 synaptic inputs are needed for neuronal
firing (Levy and
Calvert, 2021) |
|
35 |
Any set of 140 input signals arriving from random
locations on the dendritic tree can fire a neuron. Hence,
there is extreme degeneracy of input signals in firing a
neuron. A system operating by such a scheme was selected
from large number of variations since this was offering
functional advantage to the system |
Since such a scheme is expected to be used specifically,
then a possible situation must be there. If a neuron is
being held at subthreshold level by receiving nearly 130
inputs, then it needs 10 more input signals for its firing.
If only a specific cue stimulus in a specific context can
provide a specific set of input signals for 10 additional
input signals, then this possibility can be tested. |
Islet of inter-LINKed spines can provide an opportunity to
pool all the potentials at one location from where it can be
delivered in a summated manner. Dendritic spikes can be
viewed resulting from it. These can reach the axonal hillock
efficiently to cause neuronal firing for motor effect.
Inter-LINKing with spines that receive different
neurotransmitters at the islets can regulate these islets (Vadakkan, 2016a) |
|
36 |
Many
neurons are
being held at sub-threshold activation state.
|
By holding a neuron at a certain
potential below the threshold, it is possible to regulate
the neuronal output conditional upon arrival of certain
number of inputs. If these inputs can be made conditions for
an output, then it has operational significance |
Several neurons are being held at subthreshold activation
states (Seong et al., 2014). At the islets of inter-LINKed spines summation of
potentials from certain set of inputs can be guided to
generate summated potentials and even spikes that can decide
motor outputs
|
|
37 |
Input signals (postsynaptic potentials) have maximum
strength at the location of their origin, which is the spine
head. As the potentials propagate, they get attenuated in
the spine neck region. Further attenuation occurs as they
propagate towards the neuronal cell body |
When signals from a stimulus
attenuate, they may not contribute to an efficient learning
mechanism. Furthermore, signals from different spines mix in
the dendrite. Hence, most likely location for a learning
mechanism that can maintain specificity until the time of
its retrieval is expected to occur at the spine head region. |
IPL occurs between spine head regions between spines that
belong to different neurons
(Vadakkan, 2010a;
2016a). |
|
38 |
Dendritic spikes occur by the summation of nearly 10 to 50
postsynaptic potentials (of the spines) at the dendritic
region |
It is necessary to explain which spines contribute to
the potentials and explain their significance. |
Though not proved, semblance hypothesis proposed that most
of the potentials that contribute to dendritic spikes
originate from spines that belong to different neurons that
for islets of inter-LINKed spines (Vadakkan, 2016a) |
|
39 |
When current is injected into the dendrites of human layer
2/3 neurons they generated repetitive trains of fast
dendritic calcium spikes, which can be independent of
somatic action potentials (Gidon et al., 2020). |
Explanation for spike is needed |
The islets of inter-LINKed spines can lead to generation of
dendritic spikes. The net potential can drain through some
of the spines depending on the several regulatory factors (Vadakkan, 2016a) |
|
40 |
|
Conventionally it is thought that dendritic spikes are efficient detectors of specific input patterns ensuring a neuronal output (action potential) (Gasparini et al., 2004). So, a source for leakage of potentials from the dendritic area other than its propagation towards the soma needs to be explained. | The islet of inter-LINKed spines (IILSs) provides a new route for the dendritic spike to propagate. Dendritic spike can backpropagate to many IILSs & propagate through their inter-LINKed spines (that offer less resistant routes) towards the dendritic trees of different neurons from which we are not recording (so we don't record the leak). |
|
41 |
Inner sensation of certain higher brain functions occurs
without any motor actions |
Either the motor action can be voluntarily suppressed or
that there are no behavioral motor actions associated with
it |
The apical
dendrites in human layer 5 neurons are electrically isolated
from that of the somatic compartment, possibly having
independent operations of islets of inter-LINKed spines at
those remote dendritic regions
(Beaulieu-Laroche et al., 2018) |
|
42 |
When two
differential electrodes are placed at 2 extracellular
locations, extracellular potentials can be recorded. They
show oscillations. Brain operates only when the frequency of
these oscillations occurs within a narrow range |
While synaptic transmission provides one vector component,
something else constitutes the other vector component/s that
is/are expected to take place nearly perpendicular to the
direction of synaptic transmission. Brain functions (both
first-person & motor actions) are linked tightly to these
vector components. |
Propagation of depolarization along the IPL provides a
vector component almost perpendicular to that of the
synaptic transmission (Vadakkan, 2010a;
2013) |
|
43 |
Apical tuft regions of neurons of all the cortical
neuronal orders are anchored to the inner pial surface
resulting in overlapping of the dendritic arbors of neurons
from different orders. This resulted from a sequence of
movement of neuronal precursors during development |
Dendritic spines of neurons that belong to both the same
(mainly) and different neuronal orders overlap with each
other to serve certain functions |
Overlapping of dendrites that belong to different neurons
facilitate formation of inter-neuronal inter-spine LINKs.
Anchoring of apical tuft regions of all the cortical
neuronal orders facilitates this (Vadakkan, 2016a,
2019a) |
|
|
Following learning,
initially there is conscious retrieval of memory and
eventually this becomes
sub-conscious after repeated retrievals |
The process by which repeated retrieval of a
memory to a sub-conscious level must be able to explain
a framework of a mechanism of consciousness |
Explained by the semblance hypothesis
(Vadakkan, 2010b; 2019a).
Once the retrieval of memories of a certain item becomes a
routine, the inner sensations evoked by their IPL
reactivations will be added on to the inner sensation of
consciousness. Hence, such inner sensations will not be
notice individually.
|
|
|
Experimental finding of long-term
potentiation (LTP) has
shown
several correlations with behavioral motor actions that are surrogate markers of
memory retrieval |
It must be possible to explain how cellular changes during
LTP induction and learning are correlated & how this is
related to the ability for memory retrieval |
Explained by the semblance hypothesis (Vadakkan, 2019b) Foot note2 |
|
46 |
Learning takes place in milliseconds, whereas LTP
induction takes at least 20 to 30 seconds and even more a
minute |
Cellular changes during learning are expected to
get scaled-up during LTP induction in a time-dependent
manner. Need a cellular explanation for this |
Explained by the semblance hypothesis (Vadakkan, 2019b). |
|
|
Blockers of membrane fusion blocks LTP |
Need to
explain the cellular location where they act and explain how it blocks
LTP.
|
Explained by the semblance hypothesis
(Vadakkan, 2019b). |
|
|
CA2 area of hippocampus is resistant
to LTP induction.
Induction of LTP
here
becomes
possible by the removal of the peri-neural net proteins
chemically |
Cellular
mechanism responsible for LTP induction must be able to
explain this. Perineural area is involved in the cellular
mechanism of LTP induction |
Explained by the semblance hypothesis
(Vadakkan, 2019b). |
|
49 |
Several seizures spread laterally to adjacent cortices. |
Cellular mechanism responsible for seizures should be
capable of spreading laterally. |
Explained by the semblance hypothesis (Vadakkan, 2016d). |
|
50 |
Several seizures are associated with hallucinations. |
Laterally spreading seizure mechanism should be able to
explain how internal sensation of certain stimuli occur
|
Explained by the semblance hypothesis (Vadakkan, 2016d) |
|
51 |
Amyotrophic lateral sclerosis (ALS) pathology spreads laterally. |
Some structural aspects of the normal operational
mechanism aid in the lateral spread of neurodegenerative
changes under pathological conditions |
Explained by the semblance hypothesis (Vadakkan, 2016c) |
|
52 |
Relationship
between LTP, kindling, and seizures |
It is reasonable to infer that a structure-function-pathology
relationship exists that can provide
interconnecting explanations |
Explained by the semblance hypothesis (Vadakkan, 2016d) |
|
53 |
Transfer of injected dye from one CA1 neuron to the
neighboring CA1 neurons is observed in animal models of
seizures (Colling et al., Brain Res. 1996) |
Need an explanation for a physical connection between
two CA1 neurons through which dye can diffuse |
Explained by the semblance hypothesis (Vadakkan, 2016d). |
|
54 |
Loss of
dendritic spines after kindling |
An interconnected explanation for kindling, seizures and
LTP must also be able to be extended to explain loss of
spines after kindling. |
Explained by the semblance hypothesis
(Vadakkan, 2016d) |
|
55 |
CA2 area of hippocampus is resistant to seizures |
In row 44, we saw that CA2 region is resistant to LTP
induction. Hence, whatever causes resistance to LTP
induction must also be causing resistance to seizure
generation. |
Explained by the semblance hypothesis
(Vadakkan, 2016d) |
|
|
Seizures and memory
loss
are
caused by herpes
simplex viral (HSV) encephalitis |
Mechanistic explanation for both these features is expected to provide
some information about the relationship between these
findings in HSV encephalitis. |
Explained by the semblance hypothesis
(Vadakkan, 2016d). |
|
57 |
Anesthetic agents alleviate seizures. |
Mechanism of action of anesthetic agents should be able
to explain how seizure generation and propagation are
stopped by anesthetic agents. |
Explained by the semblance hypothesis
(Vadakkan, 2016d) |
|
58 |
Cognitive impairment in patients with seizure disorders. |
Mechanism of learning, memory retrieval and behavioral
motor actions are expected to be affected by the mechanism
of seizures |
Explained by the semblance hypothesis
(Vadakkan, 2016d) |
|
59 |
Intracellular electrophysiological correlate of
epileptiform activity is paroxysmal depolarizing shift
(PDS), which is a giant excitatory postsynaptic potential
(EPSP) |
A mechanistic explanation is needed for generation of a
giant EPSP at the dendritic spine area during a seizure. It
has a propensity to propagate laterally to other cortical
regions. Need a mechanistic explanation. |
Explained by the semblance hypothesis
(Vadakkan, 2016d) |
|
60 |
Neurodegenerative disorders show loss of spines and neuronal death |
An explanation is needed for contiguous spread of pathology
leading to spine loss and neuronal death.
Causative factors should be acting at specific locations to explain all
its features |
Explained by the semblance hypothesis
(Vadakkan, 2016c) |
|
61 |
Dementia in
neurodegenerative disorders |
Need an explanation for
the role of spines
in both generation of inner
sensation of memory
along with concurrent behavioral motor activity.
|
Explained by the semblance hypothesis
(Vadakkan, 2016c). |
|
62 |
Perception as
a first-person inner sensation |
A variant or a modification of the mechanism of induction of inner sensation for memory
should be able to
explain
first-person inner sensation of
perception |
Explained by the semblance hypothesis
(Vadakkan, 2015b). |
|
63 |
Apparent location of the percept different
than its
actual location |
Matching explanations using the mechanism of induction of units of inner sensation
are needed. |
The inner sensation of percept is
generated by integral of all the perceptons. Hence, the
actual location of an object need not necessarily match the
percept. This becomes clear when there is a medium that
shift the patch of light towards the eye (Vadakkan, 2015b) |
|
64 |
Homogeneity
in the percept for stimuli above the flicker fusion
frequency.
|
A mechanism for fusion of inner sensation of continuous perception of a
source of light that is affected by frequency of flickers is
needed |
Since perceptons from IPLs located at different regions in
response to one flicker has a temporal pattern of
generation, overlapping formation of perceptons from
consecutive flickers overlap and generate a continuous
percept
(Vadakkan, 2015b) |
|
65 |
Perception of object borders. |
A mechanistic explanation for the formation of first-person percept for
object borders is needed |
Only stimuli from within the border region reaches the
brain. When perceptons formed from these stimuli integrate,
they generate inner sensation of percept to generate
boarder. Similarly, stimuli from outside the borders also do
the same to generate a contrasting border of the background
(Vadakkan, 2015b) |
|
66 |
First-person inner sensation of pressure
phosphenes. |
Mechanism of generation of first-person inner sensations
is expected to provide an explanation for phosphenes
triggered by pressure over the eyeball |
Stimulation of sensory paths anywhere
along it before reaching the locations of their convergence
can lead to reactivation of IPLs for generation of
perceptons (Vadakkan, 2015b). |
|
67 |
Orientation tuning of a population of neurons in V1 before
and after training on a visuomotor task showed different
sets of neurons responding
(Failor et al., 2021) |
Neurons that fire as a consequence of associative
learning changes in the primary visual cortex varies with
time |
Based on the semblance hypothesis, the primary mechanism
of perception is not through the firing of a specific set of visual cortical neurons.
Instead, perceptons are generated at the inter-LINKed spines
on either side of an IPL
(Vadakkan, 2015b) |
|
68 |
Flash-lag
effect - When a flash
is briefly presented in a specific location adjacent to the
path of a uniformly moving object, the former is perceived
to lag the latter |
Matching explanation using the
mechanism of induction of units of inner sensation
is needed. Needs to explain how perception is affected by
relative time of arrival of a stimulus |
Explained based on the semblance hypothesis
(Vadakkan, 2022). Visual pathway has synapses that cause synaptic delay.
Overlapping reactivation of IPLs by continuous arrival of
stimuli maintains perception; whereas a fresh stimulus
undergo delay to initiate perception |
|
69 |
Inner sensation of
consciousness |
Presence of a continuous operational
mechanism for the generation of inner sensations that
depends on/contributes to maintaining the
frequency of oscillating
extracellular potentials
in a narrow range. The combined inner sensation is expected
to generate inner sense of being conscious.
|
There is a baseline oscillating
extracellular potentials as recorded by EEG. This shows
testable propagation of potentials along many IPLs
contributing to its horizontal component. Net inner
sensation generated by reactivation of inter-LINKed spines
during background state can contribute to inner sensation of
consciousness (Vadakkan, 2010b). |
|
70 |
Loss of
consciousness by anesthetic agents |
Properties of anesthetic
agents
should be able to explain how the proposed mechanism of consciousness
can be altered |
Explained by rapid chain formation of
large number of non-specific IPLs (Vadakkan, 2015a) |
|
|
Loss of consciousness during a generalized seizure and its reversal after seizure |
Mechanism of seizure generation
should be able to explain how inner sensation of
consciousness is lost.
|
Explained based on the semblance
hypothesis
(Vadakkan, 2016d). Rapid chain
formation of large number of non-specific IPLs due to
changes in ECM properties (e.g. Very low serum Na) or due to
increased excitability of neurons. |
|
72 |
Changes in consciousness with variations in the frequency
of oscillating extracellular potentials beyond a narrow
range |
Need an explanation how a narrow range of frequency of
oscillating extracellular potentials is associated with
normal state of normal state of consciousness. |
Explained based on the semblance hypothesis
(Vadakkan 2010b; 2015a).
Unconscious states show large variations in the frequencies
of extracellular potentials recordered from skull surface in
EEG (Rusalova, 2006). |
|
73 |
Effect of
dopamine in augmenting anesthetic action |
Explain a mechanism how dopamine augments anesthetic
action.
This explanation must match with the explanation for the
action of dopamine in augmenting learning (see row 11) |
Explained based on the semblance
hypothesis (Vadakkan, 2015a). Since dopamine can cause spine
expansion
(Yagishita et al., 2014), it will augment non-specific IPL formation by
anesthetic agents. |
|
74 |
Phantom sensation or pain |
Explain a mechanism for the inner sensation of pain from
a lost limb at the time of phantom sensation or phantom pain.
|
As long as the IPLs that have
received inputs from a limb remains stable in the brain, any
reactivation of this by stimuli arriving to this IPL through
a different sensory input can evoke semblance of phantom
limb or pain |
|
75 |
Referred pain |
Explain a mechanism for the inner sensation of pain from
a location different from the location where the cause of
pain is present |
Inputs two different locations
converge into one IPL at a higher neuronal order region can
lead to semblance of sensory input towards those regions (Vadakkan, 2010a,
2013) |
|
76 |
Mechanism for
innate behavior that enables survival. |
A mechanism evolving from heritable changes to explain innate behavior in
response to a stimulus |
Explained based on the semblance hypothesis
(Vadakkan, 2020).
Convergence of sensory stimuli having
different velocities is programmed in the genetic code and
executed during development that favor the formation of IPLs. |
|
77 |
Presence of a
comparable
circuitry in
a remote
animal
species explains universal nature
of a biological mechanism. |
Comparable features that show relationship of a mechanism that induces units of inner sensation using synaptically-connected neuronal circuitry among different species of animals. |
Organization of neuronal connections suggesting the
presence of a comparable IPL circuitry in Drosophila
olfactory system is explained
(Vadakkan, 2015b) |
|
78 |
Neurodegeneration resulting from repeated general anesthesia
(Baranov et
al., 2009) |
Need an explanation why the repeated induction of a mechanism of loss of consciousness
by anesthetics
can lead to
loss of spines and
eventual loss of neurons. |
Explained based on the semblance
hypothesis
(Vadakkan, 2015a).
Conversion of IPLs to inter-neuronal
interspine fusion leading to degeneration is a testable
mechanism |
|
79 |
More years of education
(increased number of associative learning events) reduces
dementia risk
(Maccora
et al., 2020) |
Should be able to explain whether redundant learning-induced
changes get induced by prolonged learning events |
Explained based on the semblance
hypothesis
(Vadakkan, 2013;
2019a). Redundant IPLs form
during different learning events as new neurons get inserted
into the circuit. |
|
|
Specific brain regions appear to be associated with specific
functions based on the lesions/ lesion studies |
These are most likely locations
of converging fiber tracts or converging locations of
specific input signals responsible for those functions |
It was possible to induce
long-term potentiation (LTP) of different strengths from
different locations of convergence inputs. Hippocampus
having convergence of all the sensory inputs has shown
maximum strength of LTP |
|
81 |
Astrocytic pedocytes cover less than 50% of peri-synaptic
area in nearly 60% of the synapses in the CA1 region of
hippocampus
(Ventura and Harris, 1999) |
Hippocampal mechanism of learning & memory must explain
the suitability of distribution of astrocytic processes.
|
Explained based on the semblance hypothesis
(Vadakkan, 2019a).
Remaining free area of the spines
favor inter-neuronal inter-spine interactions that form IPLs. |
|
82 |
Present nervous systems have
evolved over millions of years and are also the results of
certain accidental coincidences |
It is expected to become
possible to explain how the circuitry that provides all the
features can be evolved through simple steps of variations
and selection |
Explained based on the semblance hypothesis
(Vadakkan, 2020).
Sparking of the first-person inner
sensations of all the features of an item in the environment
on arrival of the fastest or first sensory stimulus from
that item started providing survival advantage to animals in
a predator-prey environment |
|
83 |
Dye diffuses from one
neuronal cell to another as the cortical neurons move from
periventricular region towards their destination indicating
formation of an inter-cellular fusion pore (Bittman
et al., 1997).
This is followed by death of nearly 70% of these cells and
survival of the remaining 30% cells.
|
It is expected to become possible to explain how an event of
inter-cellular fusion leads to selection of variants that
prevent further inter-cellular fusion. Since neurons cannot
divide further, a transient stage of fusion is expected to
trigger fusion prevention mechanism in the surviving
neuronal cells. It is also necessary to explain whether this
mechanism has any role in the unique functional property of
generation of first-person inner sensations in the nervous
system. |
Explained based on the semblance hypothesis
(Vadakkan, 2020). Dye diffusion indicates formation of fusion pores
between neuronal cells. The first occurrence of
inter-neuronal fusion is likely due to changes in membrane
composition or lack of checkpoint mechanisms to arrest
hemifusion from progressing to fusion.
|
|
84 |
Following the above stage where dye diffusion is observed,
significant neuronal death (70%) (Blaschke
et al., 1996)
and spine loss (13 to 20%) are observed |
There is a high probability that the surviving cells
have acquires an adaptation |
Explained based on the semblance hypothesis
(Vadakkan, 2020).
Following death of 70% cells, an
adaptation occurring in the surviving
cells most likely prevents any future coupling between
neurons that may result in inter-neuronal fusion. This
adaptation is suitable for maintaining IPLs for generating
useful functions. |
|
85 |
Higher brain functions take place in a narrow range of
frequency of oscillating extracellular potentials as
evidenced by EEG
(Rusalova, 2006) |
a) Both the mechanism for learning and memory retrieval
contributes vector components of the oscillating
extracellular potentials. b) The specific mechanism for both
learning and memory retrieval depends on the frequency of
oscillating extracellular potentials. |
Explained based on the semblance hypothesis
(Vadakkan, 2010a;
2013) |
|
86 |
Artificial triggering of spikes in one neuron in the
cortex causes spikes in a group of neighboring neurons in
the same neuronal order
located at short distance (25–70µm) from the stimulated
neuron
(Chettih
& Harvey, 2019). |
It should be possible to explain a mechanism that can
lead to lateral spread of firing between neurons of the same
neuronal order within a short radius. Need an explanation
for a mechanism through a path other than trans-synaptic
route. |
One explanation is propagation of depolarization across the
IPLs between spines that belong to different neurons (Vadakkan, 2013). This also explains why only
sparsely located neurons get fired, correlated in
time. |
|
87 |
The protein complexin blocks SNARE-mediated fusion by
arresting the intermediate stage of hemifusion. Complexin is
present in the spines. But docked vesicles are not found
inside the spines (in contrast to what is observed in the
presynaptic terminal). This leaves the question, "Which
inter-membrane fusion is getting arrested by complexin?" |
It is necessary to explain an inter-membrane fusion
process that can be mediated by SNARE proteins and blocked
by complexin by arresting the process at or before the
intermediate stage of hemifusion in the spines.
|
SNARE proteins provide energy for bringing together
membranes against repulsive charges and overcoming energy
barrier between abutted membranes (Oelkers
et al., 2016).
They also generate force to pull together abutted membranes
as tightly as possible (Hernandez
et al., 2012).
By initiating the fusion process by supplying energy (Jahn
and Scheller, 2006),
SNARE proteins can lead to the formation of characteristic
hemifusion intermediates (Lu
et al., 2005;
Giraudo et al., 2005;
Liu et al., 2008).
Protein complexin present within the postsynaptic terminals
(Ahmad
et al., 2012)
is known to interact with the neuronal SNARE core complex to
arrest fusion at the stage of hemifusion (Schaub
et al., 2006). |
|
88 |
Transcriptomic analyses show heterogeneity of even adjacent
neurons of the same type in the cortex (Kamme et al., 2003). |
This indicates that any mixing of the contents between these
neurons is fatal to them. Hence, there will be a robust
mechanism to prevent intercellular fusion.
|
Different mRNA profiles of adjacent neurons of even the same
type indicate that any cytoplasmic content mixing will lead
to homeostatic mechanisms such as spine or neuronal loss to
prevent it (Vadakkan, 2016c).
Ultimate purpose of it is to restrict structural aspect of
IPLs to inter-membrane hemifusion.
|
|
89 |
Heterogeneity in clinical features and pathological changes
in Alzheimer's disease (& other neurodegenerative disorders) |
1. Many factors are likely involved in the operational mechanism. 2. There will be a universal mechanism that involves different neuronal types. Failure of any of these can explain heterogeneity. |
A common
mechanism is pathological conversion of normal maximum limit
of hemifusion to pathological fusion. Clinical features
depend on a) locations of IPL fusion that can damage spines
& neurons, and b) formation of non-specific IPLs at
different locations (Vadakkan, 2016c) |
|
9 |
In excitatory neurons, spine depolarization can occur even without dendritic depolarization (Beaulieu-Laroche et al., 2018a; Beaulieu-Laroche et al., 2018b). |
Why did such a mechanism get selected? What is the
functional significance of depolarization of the spine head?
Is there any link between depolarization of the spine heads,
oscillating extracellular potentials & different brain
functions?
|
IPL
mechanism that generates units of inner sensation needs only
depolarization of spines. Lack of firing of the postsynaptic
neuron will lead to lack of motor output while units of
inner sensation occur at an inter-LINKed spine (Vadakkan, 2013;
2019a) |
|
91 |
The histological features of amyloid (senile) plaques and
neurofibrillary tangles observed in normal aging (Anderton, 1997) are also the pathological features
in Alzheimer's disease & several other disorders in the
spectrum of neurodegenerative disorders |
A mechanistic explanation for how & why intracellular
neurofibrillary tangles & extracellular plaques that are key
pathological features in neurodegenerative disorders are
observed in normal aging (but without symptoms).
|
The last stage of IPL formation is hemifusion, which is an
intermediate stage of fusion. Various factors such as viral
fusion proteins and membrane compositional changes can
overcome the check point mechanisms converging IPLs to
inter-neuronal inter-spine fusion. This will cause
cytoplasmic content mixing. Since expression profiles of
even adjacent neurons of same type are different, it leads
to neuronal damage
(Vadakkan 2016c) |
|
92 |
Therapeutic agents developed for treating seemingly
unrelated neurological diseases such as seizure disorders,
Parkinson's disease, spasticity, and hallucinations can
alleviate different headache pains |
Explanations for mechanisms of different disorders & the
operational mechanism of the system should provide
interconnected explanations for the effectiveness of
therapeutic agents in different headaches. |
By inhibiting voltage-gated sodium channels, it reduces
neuronal excitability & prevent rapid IPL formation
preventing seizures, prevents IPL formation between spines
of spiny neurons of basal ganglia, reduce inputs via IPLs to
upper motor neurons reducing spasticity, reverse/inhibit
IPLs inhibiting/reducing inner sensation of headache pains. |
|
93 |
Since
learning is expected to generate certain new circuit
connections, the circuit elements (like on a printed circuit
board) must remain separate from each other |
Properties of both neuronal membranes and extracellular
matrix should match with the new circuit connections,
functional properties imparted by them and their reversal |
Even though extracellular matrix space seems negligible between the membranes, hydration layer between the lipid membranes shows high energy barrier in artificial systems (Rand and Parsegian, 1984; Martens and McMahon, 2008; Harrison, 2015). |
|
94 |
"Representational drift" - meaning that when a brain function is repeated, set of neurons that fire changes with time (Schoonover et al., 2021; Marks& Goard, 2021; Deitch et al., 2021). |
In the case of memory, it is necessary to show redundancy in its operational mechanism, presence of a common integration mechanism and shift in the locations from where function occurs. |
Correlation between a brain function and neuronal firing
will be true for those neurons that are being held at
sub-threshold activation state and receive additional
potentials through the same IPLs.
Subthreshold activation state of a neuron can be affected by
several factors. Additional learning events can lead to the
formation of new IPLs. These can change the set of neurons
that fire (Vadakkan, 2019a) |
|
95 |
The controversial views (pdf) expressed by Camillo Golgi against Ramón y Cajal's
interpretations of results obtained from modified Golgi
staining protocols |
The chemistry behind the modification of original Golgi staining protocol must be able to provide reasons for this controversy. Such an explanation is expected to become possible when we understand the operational mechanism of the brain. |
Spines within the islets of inter-LINKed spines are
connected via an oxidation state dependent manner (Vadakkan, 2022) |
|
96 |
Formation of new neurons in the hippocampus, especially in non-stationary environments. |
The operational mechanism should be able to explain
functional advantage provided by insertion of new neurons |
Both input and output connections of new neurons will
continuously alter the existing circuitry. Repetition of
same associative learning will make new IPLs at higher
neuronal orders increasing number of sparse storage
mechanisms (Vadakkan, 2011a) |
|
97 |
Loss of spines and formation of new spines during learning
(Frank et al., 2018)
|
There is a mechanism that leads to loss of spines during learning. Formation of new spines should accomplish something new that can facilitate further learning. |
The last stage of permitted intermembrane interaction leading to IPL formation is inter-membrane hemifusion, which is an intermediate stage of membrane fusion. Several factors can overcome the checkpoint needed to arrest the changes at the stage of hemifusion. |
|
98 |
Generalization is seen in the Transformers of the large
language models (LLMs) that use neural networks organized
using a hidden layer |
A mechanistic explanation is needed to explain where and
how the hidden layer operates. This must explain how the
system generates an appropriate output in response to a new
prompt that the system never got exposed in the past |
Basic concept of “islet of inter-LINKed spines” in the
semblance hypothesis matches with that of the “attention heads” in the
Transformers of LLMs (Vadakkan, 2024) |
| 99 |
|
Reasons are necessary to provide explanation for high frequency of oscillations in layers 2 &3 & low frequency in deeper layers | Apical tuft region of neurons of all the cortical layers are attached to the marginal zone close to the pial layer (innermost covering layer of the cortex). Layer 1 cortical neurons that are mostly GABAergic send horizontal processes interconnecting several postsynaptic terminals of apical tufts. Hence, dendritic arbors of neurons of all the cortical layers overlap maximally in layer 2, followed by layers 3, 4, 5, and 6 in decreasing order. This increases the probability of formation of maximum number of synapses (vertical component) & inter-neuronal inter-spine interactions (horizontal component) in layer 2. This in turn explains high frequency of oscillations of potentials across neurons that are connected vertically (through synapses) & horizontally (through IPLs) in a decreasing order from layer 2 to 6 (Vadakkan, 2015). |
|
IPL: Inter-postsynaptic functional LINK
Foot note1
If we provide a set of colors and picture against each color
and ask people to learn the association in one second, most
people will be able to learn two or more associations
during this period. This means that associative learning can
take place in milliseconds. What type of a change can occur
in less than one second?
Foot note2
Around 30 specific experimental findings are related to
the association between LTP, ability to learn, behavioral
motor action at the of memory retrieval and biochemical
changes following learning. It was possible to satisfy all
the constraints arising from those
findings in an interconnected manner. |
|||
Table
2.
Features
of the system from different levels
that need
to be
explained independently and by an
inter-connectable
manner using a derived solution
A complete understanding of the operational mechanism leading to the first-person properties will only be achieved by carrying out the gold standard test of its replication in engineered systems. Even though replication of motor activities to produce behaviorally equivalent machines may seem adequate, the work will not be complete until first-person properties of the mind are understood. Engineering challenges with this approach include devising methods to convert the first-person accessible internal sensations to appropriate readouts. Experiments to translate theoretically feasible mechanisms of its formation both by computational and engineering methods are required. Feasibility to explain various brain functions both from first-person and third-person perspectives qualifies it as a testable hypothesis. Research findings from different laboratories have been examined in terms of the semblance hypothesis. The present work resulted from curiosity to understand the order behind the seemingly complex brain functions. In this attempt, I have used some freedom to seek a new basic principle in order to put the pieces of the puzzle together. This work wouldn't have become possible without a large amount of research work painstakingly carried out by many researchers over many years. Even though the present hypothesis is compatible with experimental data, it must be considered unproven until further verification of its testable predictions are made.
Video presentations
1.
A
testable hypothesis of brain functions
2. How to study inner sensations? Examples from mathematics
4.
List of third-person findings and the derivation of the solution for
the nervous system
6.
Induction of units of inner sensation
8. Potentential mechanism for neurodegeneration
9. LTP: An explanation by semblance hypothesis
--------------------------------------------------------------------------------------------------------------------
The challenge: "What I cannot create (replicate), I do not understand" – Richard Feynman. The rigor with which we should try to solve the nervous system must be with an intention to replicate its mechanism in an engineered system. Everything else will follow.
The reality: We are being challenged to find a scientific method to study the unique function of the nervous system - how different inner sensations are being generated in the brain concurrent with different third-person observed findings. We cannot directly study them using biological systems. But we can use all the observations to try to solve the system theoretically, followed by verifying its predictions.
The optimism:“What are the real conditions that the solution must satisfy?” If we can get that right, then we can try and figure out what the solution is" – Murray Gell–Mann
The expectation:
We are
likely able to solve
the mechanism of the nervous system functions
in multiple
steps.
First, using constraints offered by all the observations, it is necessary
to
derive a solution (most likely a first principle) that can unify those observations.
This can be followed by further verifications by triangulation methods
and examining comparable circuitries in different animal species.
Once identified
and verified,
we can expect to
replicate the
mechanism
in
engineered systems.
The hope: We will give everything we can. Together we will explore it!